

Ridhima Jain – ridjain.14@gmail.com
August 7th, 2025
Edited by the YNPS Publications team.
1. Abstract
Genetic ancestry is a tool that constitutes genetic information about the people one has descended from, tracing their genetic relationships. Traditionally used to identify geographical origins, ancestral roots, and criminal suspects, genetic analysis has recently transformed Alzheimer’s research: studies have revealed that apolipoprotein E (APOE) is influenced by genetic ancestry, with the risk and severity of the disease potentially varying based on the geographic background of the individual who inherits the gene. This essay encompasses the contribution of genetic ancestry in understanding the genetic and molecular behavior of APOE and how its subsequent practice could sustain future solutions in preventing Alzheimer’s disease (AD). By examining the work of Griswold, Anthony J., et al. (2017) for the transcriptional variation in APOEɛ4 between African American local ancestry (ALA) and European local ancestry (ELA), including the finding of A1 reactive astrocytes prevalence in ELA brain tissue and its relation to APOEɛ4 characterization, and Le Guen, Yann, et al. (2023) for the discovery of the R145C variant in the APOEɛ3 and its exacerbation of AD within the ɛ3/ɛ4 genotype in African ancestry, we observed genetic ancestry’s capability to facilitate patterns in gene expression, becoming a focal reference in determining the risk of disease exposure amongst different geographic populations. These results suggest that comparing the genetic ancestry between diverse geographic communities can aid in deciphering the extensive profile of APOE and ultimately strategize AD prevention methods.
2.1 Local Ancestry and Gene Expression
Though the cause behind AD remains an enigma, over the years, genetics has revealed that the presence of the apolipoprotein E plays an integral risk factor in promoting AD. APOE is an inherited protein that aids in transporting cholesterol and fat into the bloodstream and yields three allele variants, each attributed with a different effect on AD risk: APOEɛ2 can delay Alzheimer’s; APOEɛ3 is the most common variant, hosting a neutral effect on the disease; APOEɛ4 increases the risk of early-onset Alzheimer’s. However, the impact of each allele varies with local genomic ancestral (LA) region: if one has inherited the APOEɛ4 allele from an African ancestor via ALA, they have the African risk of AD from APOEɛ4; the same follows for APOEɛ4 inheritance through ELA. Since no trending differences in the APOEɛ4 mRNA were observed between AA and ELA, Griswold, Anthony J., et al. (2017) deduced that the shielding effect of the protein must be located in the non-coding variants and hypothesized that they could impact the genetic expression engendering the risk variation. Thus, Griswold, Anthony J., et al. (2017) focused on recognizing distinctions in gene expression of APOEɛ4 in both LAs. To achieve this, the study procured samples of brain tissue from ALA and ELA AD patients who are all homozygous carriers of the APOEɛ4 gene. Additionally, the employment of single-nucleus RNA-sequencing (snRNA-seq) ensures that the bulk transcriptome—the average genes expressed—allows abundantly unbiased expression profiles to be analyzed.
2.2 Investigating ALA Missense Variants on AD Risk
With APOEɛ4 and ɛ2 surging AD risk within ELA, insufficient information stands on how the APOE gene catalyzes late-onset Alzheimer’s in African ancestry. Although a few missense variants have been identified within individuals of European ancestry, for instance, the Val236Glu (V236E) and Arg251Gly (R251G), which are concerned with inhibiting AD susceptibility, the scarcity in research about missense variants is due to their prevalence in ALA and their lack of representation in AD genetic studies. Moreover, 2 variants with minor allele frequencies were exclusively discovered in APOE transcripts belonging to ALA: Arg145Cys (R145C) and Arg150His (R150H), the former more common with a minor allele frequency of roughly 2% and the latter with approximately 0.1%. Both variants are located amid the receptor-binding site at amino acid residues 136-150 in the N-terminal domain of APOE, and previous studies have discerned that R145C is always in accord with ɛ3 and R150H behaves similarly with ɛ2. Since the most common among these variants are always present in an African haplotype, i.e., only found in ALA, Le Guen, Yann, et al. (2023) aimed to determine if R145C and R150H influence AD risk, implicating the improvement in ancestry-unique risk prediction.
3. Methodology
All findings and analyses presented in this essay are contrived from various research databases, including PubMed, Stanford Medicine, and JAMA Network. The investigations conducted by Griswold, Anthony J., et al. (2017) and Le Guen, Yann, et al. (2023) are funded by numerous government-affiliated laboratories and facilities.
To evaluate how the gene expression profile of APOEɛ4 compared between individuals of African and European descent, Griswold, Anthony J., et al. (2017) discreetly 1) selected homozygous APOEɛ4 participants from both ancestral backgrounds; 2) employed genotyping data along with the SHAPEIT tool ver. 2 and the RFMix algorithm to genomically characterize the APOEɛ4 allele, focusing on a central LA region yielding consecutive LA blocks while also accounting for any regulatory factors; 3) isolated nuclei under the same conditions, extracting tissues with the Nuclei Isolation Kit: Nuclei EZ Prep; 4) performed snRNA-seq where a nuclei concentration of 1200 nuclei/μl was isolated at ~7,000 nuclei per sample, each barcoded with Gel Bead-in Emulsions to succor enhanced analysis; 5) utilized data integration to capture shared cell states among various samples, and identified differently expressed genes in ALA and ELA through the MAST test.
The probe concerning the relationship between the missense variants in African ancestry and AD vulnerability has been executed as a case-controlled study. Le Guen, Yann, et al. (2023) divided their study population into three cohorts: the Alzheimer’s Disease Sequencing Project (ADSP) served as the discovery sample, the Alzheimer’s Disease Genetics Consortium (ADGC) as the replication sample, and the Million Veteran Program (MVP) from the U.S. Department of Veterans Affairs as the validation sample. All participants were of African descent, with 3% identified as Nigerian and the remainder as African American. Each cohort consisted of neurotypical and AD-diagnosed individuals; both groups had their APOE genotype recorded. Multiple methods of stratified analyses were utilized to assess the AD risk association with the variants.
4. Results
This section divides the results amassed from the two studies: Griswold, Anthony J., et al. (2017) will cover the first four findings, and will proceed with the observations exposed by Le Guen, Yann, et al. (2023).
Key findings gathered from the Griswold, Anthony J., et al. (2017) study embraced the discovery of 32 different neuronal clusters varying in the proportion of cells per cluster and in APOE expression.
Significant evidence obtained from Le Guen, Yann, et al. (2023) consisted of the relationship between the AD risk and R145C in different APOE genotypes.
4.1 snRNA-seq Clusters
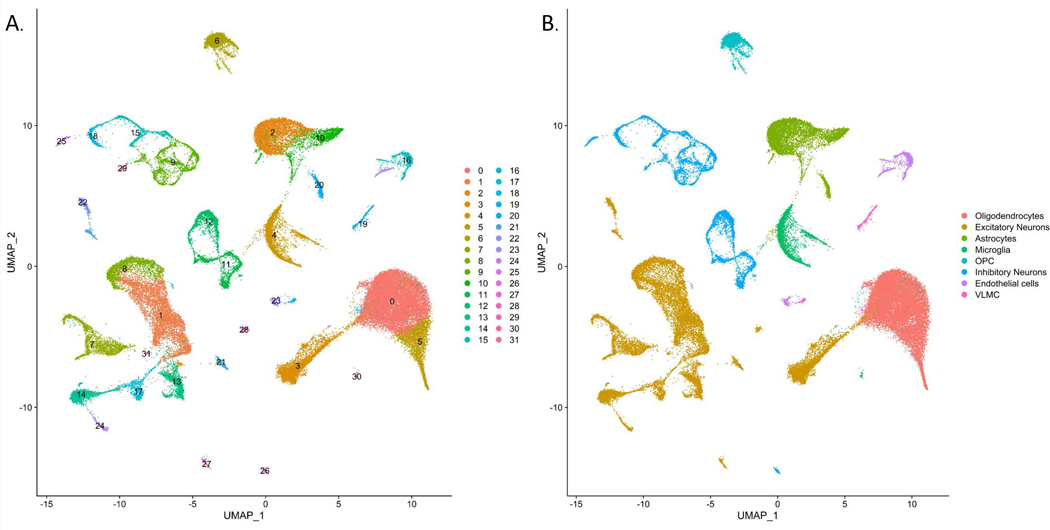
Among the 60,908 total nuclei, thirteen excitatory neuron clusters, two oligodendrocyte clusters, two microglia clusters, three astrocyte clusters, eight inhibitory neuron clusters, one oligodendrocyte precursor cell cluster, one vascular leptomeningeal cell (VLMC) cluster, and two endothelial cell clusters were observed (Figure 1). Across all eleven samples of the integrated nuclei, the excitatory neuron clusters constituted a predominant portion of the total cells (~33% of the total cells), followed by the oligodendrocyte clusters (~29% of total cells). The lowest portion of the cells was covered by the VLMC cluster (~1% of total cells).
Figure 1 (PMC)

4.2 Cluster Comparison Between ALA and ELA
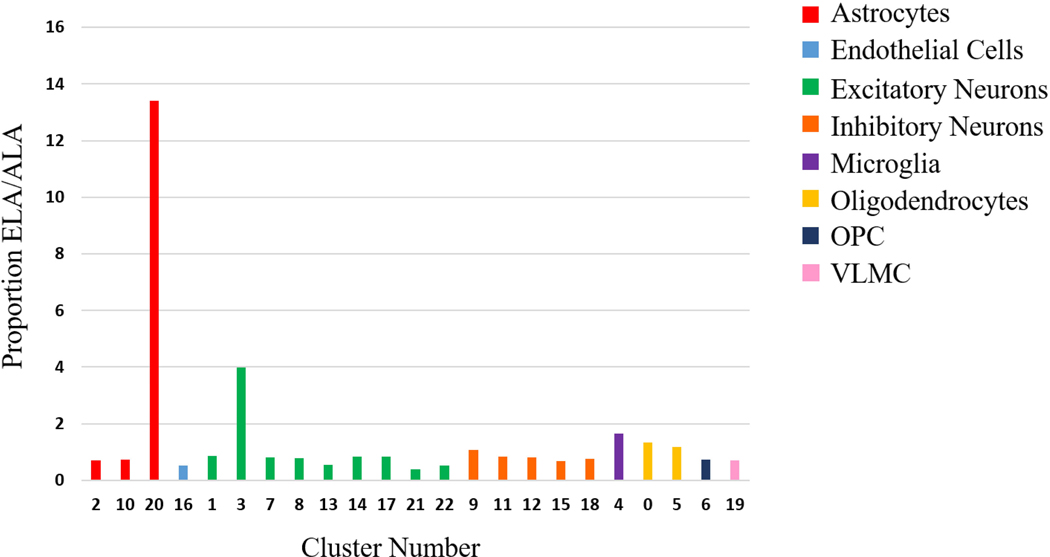
Although the proportion of cells per cluster proved relatively similar between both ancestries, noticeable distinctions were highlighted in clusters 3, 20, and 21 (Figure 2). While both excitatory neuronal clusters demonstrated a 2-fold difference between the ancestries, the proportion of total cells for cluster 20 in ELA was 13x greater than the proportion of total cells in ALA samples, despite observations mainly extracted from two ELA samples.
Figure 2 (PMC)
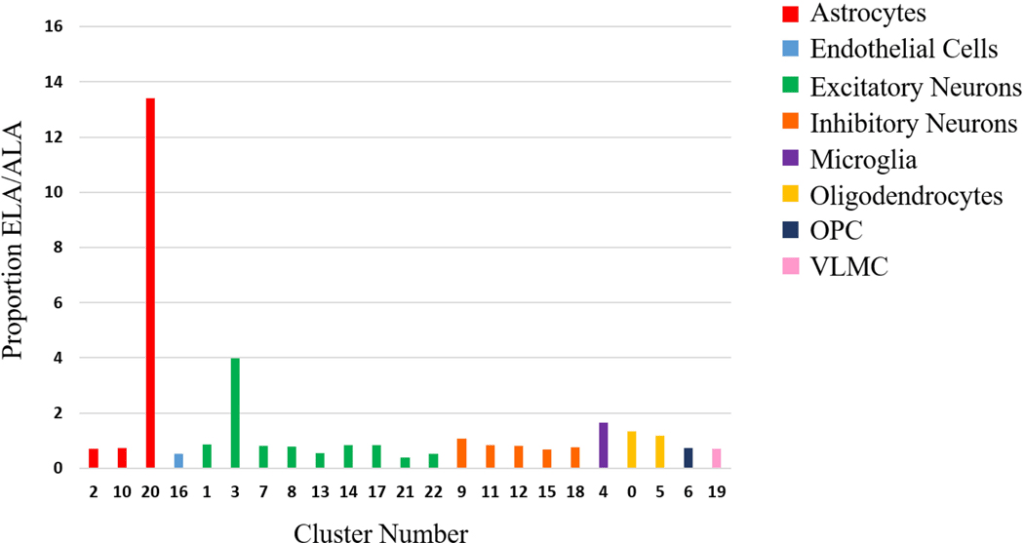
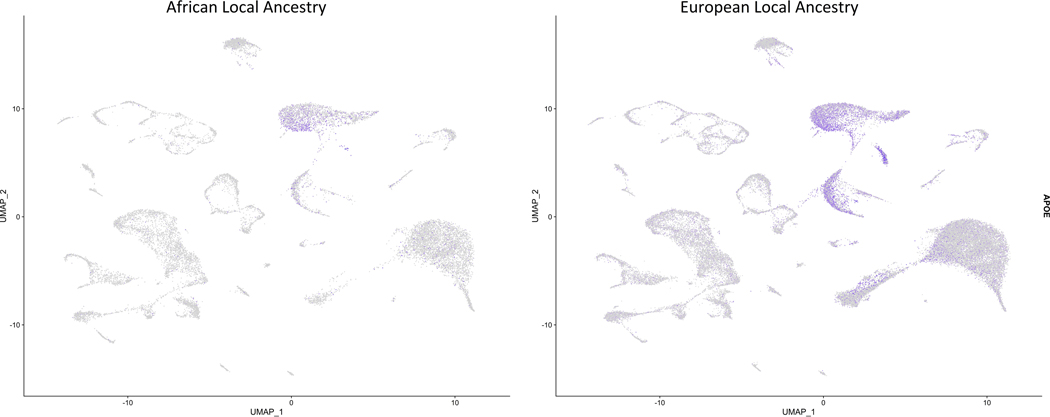
4.3 APOEɛ4 Expression
The expression of APOEɛ4 exhibited notable differential expression in ⅛ of the clusters, with all fold changes in the negative direction (Table 3). The most significant fold change was discerned in cluster 4, where ALA cells illustrated 2.2-fold fewer APOEɛ4 than ELA cells. ELA samples also yielded higher prevalence in APOEɛ4 expression than in the ALA samples (Figure 3). The association between APOEɛ4 and the ancestral groups was corroborated by a second analysis comparing three samples of each ancestry, instead, in which samples collected from the same site displayed an exceptionally higher APOEɛ4 expression in the ELA (p < 3.5E-318, FC = 1.42).
Table 3: APOE Differential Expression Analysis (PMC)
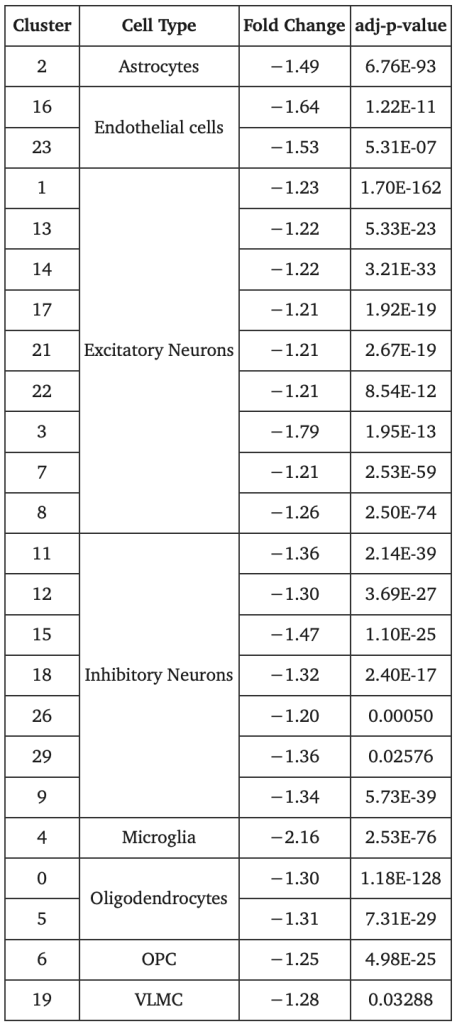
Figure 3 (PMC)

4.4 Transcriptional Examination of Astrocyte Cluster 20
To characterize cluster 20, its transcriptomic profile was compared to those of the other astrocyte clusters. The identified differentially expressed genes suggest that cluster 20 evinced indication of reactive astrocytes, overexpressing regulative proteins like VIM (p = 9.33E-267, FC = 2.09) and GFAP (p = 2.88E-192, FC = 2.11), and genes that are upregulated by A1 astrocyte reactivity in mice like OLFM1 (p = 8.17E-204, FC = 1.90) and IFITM3 (p = 1.08E-215, FC = 1.74).
4.5 Association between R145C and AD Risk in the ɛ3/ɛ4 Genotype
Stage 1 discovery datasets regarding ɛ3/ɛ4 individuals with R145C presented 52 cases and 19 controls, ascertaining increased AD susceptibility, contrasting to those without the variant (OR, 3.01; 95% Cl, 1.87-4.8; P = 6.0*10-6) (Table 3). Similarly, stage 2 delivered 23 cases and 21 control samples containing R145C (OR, 2.20; 95% Cl, 1.04-4.65; P = 0.04), and the stage 3 MVP replication with R145C present in 11 cases and 149 controls (OR, 1.90; 95% Cl, 0.99-3.64; P = 0.051). Furthermore, an association between R145C in ɛ3/ɛ4 and early-onset Alzheimer’s was identified amongst the patients across all three stages: stage 1 – β, -5.87 years; 95% Cl, -8.35 – -3.4; P = 3.4*10-6; stage 2 – β, -5.23 years; 95% Cl, -9.58 – -0.87; P = 0.02; and stage 3 – β, -10.15 years; 95% Cl, -15.66 – -4.64; P = 4.0*10-4. The ɛ2/ɛ3 and ɛ3/ɛ3 genotypes indicated no ostensible link between AD risk and the R145C variant.
Table 3: R145C Association with Alzheimer’s Disease Risk and Age at Onset (JAMA)

The OR (95% Cl) comparison for AD among the APOE genotypes additionally highlighted how the OR estimates concerning the missense carriers of ɛ3/ɛ4 overlapped with the OR estimates of those with ɛ4/ɛ4 in the stage 1 (Figure 2A) and stage 2 (Figure 2B) samples. This observance was validated after performing sensitivity analyses via 45% and 75% ancestry cutoffs; the results are independent of the ALA cutoffs and the sampling method.
Figure 2 (JAMA)

Note that R150H was omitted from further scrutiny as it demonstrated no association or significance on AD risk among all the APOE genotypes.
5. Discussion
After appraising their results, Griswold, Anthony J., et al. (2017) affirmed that the APOE gene produced the most significant differential gene expression in the LA locus. More APOEɛ4 was generated from ELA carriers of APOEɛ4 around the locus than the ALA carriers of the same allele, and, in fact, the gene was largely expressed in all the ELA groups but absent in the ALA samples, emphasizing the influence of differential gene expression in the AD risk between the two ancestries.
Additionally, the veritable cell disproportion in cluster 20 has likely been categorized as A1 reactive astrocytes. Reactive astrocytes are dichotomized as either A1 or A2: A1 are activated by neuroinflammatory conditions and are malicious to neural synapses, linked to cellular neurodegeneration; A2 are activated by poor cardiovascular health and designed to protect neurons by upregulating neurotrophic factors. Due to the atypical upregulation of GFAP, B2M, OLFM1, and IFITM3, the reactive astrocytes are deemed as A1. Although A1 reactive astrocytes are reportedly amassed within AD neural profiles and are contributive to early-onset AD, a mouse study conducted by Shi, Yang, et al. (2017) reveals that augmented APOEɛ4 can induce neuroinflammation under pathological conditions, consequently triggering toxic A1 reactive astrocytes and promoting neurodegeneration. Since cluster 20 abjectly expressed APOEɛ4, the result of cluster 20 comprising 2 ELA and no ALA is merely “the biological heterogeneity of AD” or is relevant to the high expression of APOEɛ4 in ELA, is inconclusive, and demands further exploration.
As for Le Guen, Yann, et al. (2023), APOEɛ3[R145C] is related to increased AD risk, including susceptibility to early-onset AD, within the ALA of the ɛ3/ɛ4 genotype. Despite examining a variegated community of African ancestry and focusing analyses on the APOE locus amongst all samples, the findings remained consistent and are implicated to not have been conflated by the LAs, and that R145C is the responsible variant.
R145C was observed in approximately 4% of all the African American subjects and 1% in African American individuals with the ɛ3/ɛ4 genotype. Moreover, Alzheimer’s disease (AD) aggravation was observed exclusively in individuals with the ɛ3[R145C]/ɛ4 heterozygous genotype, producing a risk profile similar to that of homozygous ɛ4 carriers. In contrast, homozygous ɛ3[R145C] individuals did not exhibit an increased risk, although the sample size was too small to draw definitive conclusions. This pattern suggests that the presence of the ɛ4 allele may negate a protective biochemical property associated with homozygous ɛ3[R145C], rather than introducing a new harmful function that exacerbates AD risk. Although the indeterminable cause behind this mechanism, it is speculated that because of R145C’s location within APOE’s receptor-binding region, it has a truncated binding affinity towards various degrees of low-density lipoprotein (LDL) receptors, which are needed to transport lipids across different cells, allowing glial cells to delete pathogenic proteins, and influencing synaptic function. In a similar study, Abou, Ziki, Maen D., et al. (2013) found that their African American samples of the R145C variant demonstrated poor binding affinity to heparan sulfate proteoglycan, prompting APOE to interact ineffectively with the LDL receptor, intimating that the variant’s behavior in APOE receptor-binding may refer to its AD pathogenicity.
Nevertheless, both Griswold, Anthony J., et al. (2017) and Le Guen, Yann, et al. (2023) acknowledge that a larger and genetically diverse sample size is necessary to comprehensively inform the assessment of AD risk, at least within African American individuals, and in understanding the constitutive and regulatory elements involved in APOE, which can succor development methods to obstruct APOE activity and alleviate AD risk within those with the gene.
6. Conclusion
The observance of A1 reactive astrocytes and its correspondence to the APOEɛ4 genomic profile between African American and European ancestry, along with the characterization of APOEɛ3[R145C]/ɛ4, empirically suggest how genetic ancestry can be mobilized to help uncover the comprehensive behavior of APOE and its effect on early-onset Alzheimer’s, and how sustaining its practice can pave the way for effective solutions to preclude AD by focusing on the genetic stimulus.
That being said, the cause explaining the difference in APOEɛ4 between both ancestries is currently unknown. While sequencing differences in both LAs could manifest this distinction, their impact on the other APOE genotypes between different ancestries requires additional study. Hence, genetic ancestry in Alzheimer’s research is still developing and needs further exploration to achieve progress against AD.
References
Abou Ziki, M. D., Strulovici-Barel, Y., Hackett, N. R., Rodriguez-Flores, J. L., Mezey, J. G., Salit, J., Radisch, S., Hollmann, C., Chouchane, L., Malek, J., Zirie, M. A., Jayyuosi, A., Gotto, A. M., & Crystal, R. G. (2013). Prevalence of the Apolipoprotein E Arg145Cys Dyslipidemia At-Risk Polymorphism in African-Derived Populations. The American Journal of Cardiology, 113(2), 302–308. https://doi.org/10.1016/j.amjcard.2013.09.021
Genetic Ancestry. (n.d.). Genome.gov. Retrieved August 7, 2025, from https://www.genome.gov/genetics-glossary/Genetic-Ancestry
goldmanb@stanford.edu, img src=’https://med stanford edu/news/media-contacts/bruce_goldman/_jcr_content/image img 620 high jpg/goldman-bruce-90 jpg’ alt=’Bruce G. B. G. B. G. is a senior science writer in the O. of C. E. him at. (2022, May 31). Scientists find genetic Alzheimer’s risk factor tied to African ancestry. News Center. https://med.stanford.edu/news/all-news/2023/02/alzheimers-genetics.html
Griswold, A. J., Celis, K., Bussies, P. L., Rajabli, F., Whitehead, P. L., Hamilton‐Nelson, K. L., Beecham, G. W., Dykxhoorn, D. M., Nuytemans, K., Wang, L., Gardner, O. K., Dorfsman, D. A., Bigio, E. H., Mesulam, M. M., Weintraub, S., Geula, C., Gearing, M., McGrath‐Martinez, E., Dalgard, C. L., & Scott, W. K. (2021). Increased APOE ε4 expression is associated with the difference in Alzheimer’s disease risk from diverse ancestral backgrounds. Alzheimer’s & Dementia, 17(7), 1179–1188. https://doi.org/10.1002/alz.12287
https://www.facebook.com/NIHAging. (2023, July 13). Genetic variant specific to African ancestry is associated with increased Alzheimer’s risk. National Institute on Aging. https://www.nia.nih.gov/news/genetic-variant-specific-african-ancestry-associated-increased-alzheimers-risk
Jorde, L. B., & Bamshad, M. J. (2020). Genetic Ancestry Testing. JAMA, 323(11), 1089. https://doi.org/10.1001/jama.2020.0517
Le Guen, Y., Raulin, A.-C., Logue, M. W., Sherva, R., Belloy, M. E., Eger, S. J., Chen, A., Kennedy, G., Kuchenbecker, L., O’Leary, J. P., Zhang, R., Merritt, V. C., Panizzon, M. S., Hauger, R. L., Gaziano, J. M., Bu, G., Thornton, T. A., Farrer, L. A., Napolioni, V., & He, Z. (2023). Association of African Ancestry–Specific APOE Missense Variant R145C With Risk of Alzheimer Disease. JAMA, 329(7), 551–560. https://doi.org/10.1001/jama.2023.0268
Li, K., Li, J., Zheng, J., & Qin, S. (2019). Reactive Astrocytes in Neurodegenerative Diseases. Aging and Disease, 10(3), 664. https://doi.org/10.14336/ad.2018.0720
Shi, Y., Yamada, K., Liddelow, S. A., Smith, S. T., Zhao, L., Luo, W., Tsai, R. M., Spina, S., Grinberg, L. T., Rojas, J. C., Gallardo, G., Wang, K., Roh, J., Robinson, G., Finn, M. B., Jiang, H., Sullivan, P. M., Baufeld, C., Wood, M. W., & Sutphen, C. (2017). ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy. Nature, 549(7673), 523–527. https://doi.org/10.1038/nature24016
Study finds differences in APOE ɛ4 expression based on genetic ancestry. (2021, July 1). National Institute on Aging. https://www.nia.nih.gov/news/study-finds-differences-apoe-e4-expression-based-genetic-ancestry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Leave a comment