

Saanvi Talwar – staffwriter.neuropsychology@gmail.com
July 29th, 2025
Edited by the YNPS Publications Team.
Abstract
Working memory (WM) is the cognitive system that stores and processes information temporarily. This is crucial to learning, reasoning, and emotion regulation, particularly in adolescence when the prefrontal cortex (PFC) is developing at a fast rate. Stress impacts WM via activation of the hypothalamic-pituitary-adrenal (HPA) axis and sympathetic nervous system (SNS), whose interaction controls PFC functioning. Sudden stress, including coordinated HPA and SNS activation, can temporarily enhance WM through the optimum catecholaminergic and glucocorticoid signaling in the PFC to augment executive processing. Dysregulated or chronic stress instead produces neurotoxicity in the form of dendritic retraction and synaptodendritic injury in the PFC and hippocampus, structural brain changes such as hippocampal volume reduction of 14–17%, and permanent WM deficits. Animal models suggest that adolescent chronic stress during early adolescence is an accelerator of a fivefold decline in WM in adulthood under acute stress conditions, reflecting long-term susceptibility. This review integrates behavioral, hormonal, neuroimaging, and developmental evidence to account for dose- and time-dependent, bidirectional functions of stress on adolescent WM with important implications for cognitive resilience-building interventions.
1. Introduction
1.1 The Significance of Working Memory and Brain Plasticity In Adolescence
Working memory is an executive function of the central executive that allows people to hold, update, and manipulate information necessary for complex cognition, including reasoning, problem-solving, and goal-directed action (Baddeley, 2012). Working memory predominantly depends on the integrity of the dorsolateral prefrontal cortex (DLPFC), an area of the brain that undergoes great structural and functional reorganization in adolescence. During this window of development, synaptic pruning strengthens neural connections and refined myelination improves processing rate and efficiency of connectivity (Casey et al., 2008). Ongoing development of fronto-parietal circuits and the coupling between them and limbic structures such as the hippocampus and amygdala provides a context of heightened neuroplasticity and vulnerability, resulting in the highly competent yet vulnerable adolescent brain. Psychosocial challenges such as academic demands, peer stressors, and family conflict are ubiquitous and potent in adolescence and tend to generate stress reactions whose impact on WM and related processes is frequently complex (Romeo, 2013).
1.2 Neuroendocrine Systems Guiding Stress Responses
Stress perception provokes the concerted activation of the two main neuroendocrine systems: the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic nervous system (SNS). Activation of the HPA axis causes the secretion of cortisol, a glucocorticoid hormone that binds to mineralocorticoid and glucocorticoid receptors with high density in the hippocampus and PFC and with strong modulatory effects on neuronal excitability and synaptic plasticity (Joëls et al., 2011). The impact of cortisol on cognition is subtle and concentration-dependent: moderate levels assist WM through the enhancement of synaptic functioning and gene expression associated with plasticity, but chronic or prolonged cortisol interferes with PFC function through receptor overactivation and downstream genomic effects. Meanwhile, the SNS mediates rapid catecholaminergic responses, releasing norepinephrine and epinephrine that stimulate adrenergic receptors in the PFC. These catecholamines control the rates of neuronal firing and the signal-to-noise ratio in essential WM neural circuits, but high levels disrupt these and impair cognitive control (Arnsten, 2009). The particular interaction and timing between HPA and SNS activations decide if and how stress promotes or degrades working memory, a connection further complicated by maturing adolescent brains.
2. Methods
This systematic review brings together evidence from main experimental and meta-analytic studies:
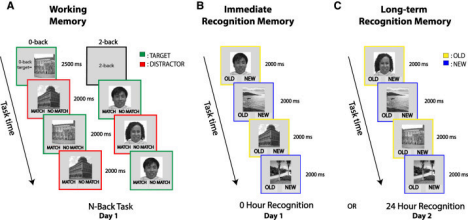
● Tsai et al. (2021) employed the Trier Social Stress Test for Children (TSST-C) to induce acute psychosocial stress in 81 children and adolescents between the ages of 8 and 15 years. The salivary cortisol and alpha-amylase levels were used as indicators of HPA and SNS activation, respectively, before and after stress induction. The task of an emotional n-back working memory was administered after stress to test cognitive performance during stress conditions.
● Shields et al. (2013) meta-analyzed 63 experiments that assessed acute stress’s effect on episodic and working memory and also explored moderators of stress timing relative to the testing of cognition, stress intensity, and participant variables.
● Chaby et al. (2015) used a chronic unpredictable stress (CUS) regimen in adolescent rats with a series of stressors for 30 days. The animals were then tested in adulthood on working memory tasks under acute novelty stress to ascertain the long-term cognitive effects of adolescent stress exposure.
● McEwen et al. (2015) consolidated neuroanatomical consequences of chronic stress in humans and animals. This focuses on changes in dendritic morphology, volumetric changes in the hippocampus and PFC, and their implications for cognitive and emotional dysfunction.
Quantitative relevant information such as levels of hormones, brain volumetric measurements, and working memory accuracy were abstracted and consolidated to construct an integrated neuropsychological model detailing stress-WM interactions during adolescence.
3. Results
3.1 Acute Stress and Working Memory: Enhancer or Impairer?
The acute stress effect on working memory is complex and extremely sensitive to the integration between the SNS and HPA axis activation. TSST-C, a thoroughly validated psychosocial stressor that activates social-evaluative threat, consistently induces increases in sympathetic biomarkers such as alpha-amylase and cortisol, and thus allows for probing of neuroendocrine effects on cognition extensively (Tsai et al., 2021). In the current study, individuals with concordant high activation of alpha-amylase and cortisol showed a notable 10–15% improvement in WM accuracy, particularly in older teenagers (13–15 years), who possessed more mature regulatory circuitry. The enhancement represents that simultaneous activation of glucocorticoid and catecholaminergic systems can optimize PFC neural processing to support executive functions needed by WM tasks. Conversely, those with conflicting results, such as high cortisol without concomitant SNS activation, exhibited up to a 20% elevation in WM error and false alarms. This indicates that stress system activation imbalance harms WM. Such findings are consistent with the “dual-system” theory of stress, in which glucocorticoid and catecholamine signaling within balance is necessary for WM modulation to function under acute stress optimally (Arnsten, 2009; Tsai et al., 2021).
Augmenting this, Shields et al.’s (2013) meta-analysis demonstrated that working memory performance is enhanced by acute stress only within a very narrow “optimal window” of stress severity and duration. Somewhat before or simultaneous with cognitive processing, moderate stress is likely to enhance WM, while higher intensity or prolonged stress exposure has a debilitating influence. This inverted-U-shaped dose-response curve highlights the delicate neurochemical balance required to facilitate cognition so that only moderate levels of norepinephrine and cortisol increase signal-to-noise ratios and synaptic plasticity in the PFC, but beyond this level elicit receptor saturation and subsequent inhibition of PFC activity.
Neurobiologically, acute stress enhances catecholamines in the PFC, augmenting stimulation of dopamine D1 receptors that strengthen task-relevant neural firing and suppress distracting stimuli, thereby supporting WM maintenance and manipulation (Arnsten, 2009). Cortisol simultaneously acts on mineralocorticoid receptors at low to moderate levels to augment synaptic potentiation and gene transcription for plasticity but at high levels to activate glucocorticoid receptors that degrade PFC function and memory processing (Joëls et al., 2011). Together, these intersections produce the transient cognitive improvement or decrement observed behaviorally.
3.2 Chronic Stress: Lasting Neuroanatomical and Cognitive Damage
On the other hand, repeated or chronic stress experienced by adolescents has far-reaching, adverse consequences on working memory primarily through neurotoxic action involving the PFC and hippocampal structure and function. Findings with animal models of chronic unpredictable stress (CUS) applied to adolescence are effective evidence. Chaby et al. (2015) demonstrated that rats subjected to 30 days of heterogeneous stressors (restraint, cold stress, social isolation) evidenced a fivefold reduction in the accuracy of working memory when tested with acute novelty stress in adulthood relative to unstressed controls (p < 0.001). This staggering impairment indicates that chronic adolescent stress sensitizes WM circuits to increased vulnerability, so subsequent acute stressors elicit exaggerated cognitive deficits. Notably, associative and reference memory domains were spared, emphasizing the specificity of WM-related neural circuits, and most prominently those incorporating the PFC, to chronic stress-induced damage. This “double-hit” finding captures how early-life stress imposes indelible marks upon neural circuitry, making people vulnerable to cognitive interference later in life.
Human neuroimaging and neuroendocrine studies validate such findings in animals. Chronic stress has been linked with alterations in brain structure, including 14–17% reductions in hippocampal volume (Lupien et al., 2009, cited in McEwen et al., 2015) and extensive dendritic loss in PFC pyramidal neurons, with reductions in dendritic spine density of approximately 30–40%. Such neuroanatomy undermines synaptic connectivity and the flexibility of the PFC’s executive control and working memory maintenance capability. Furthermore, chronic stress induces hypertrophy and increased excitability of the amygdala, further increasing emotional interference in cognitive performances, which also impairs WM performance (McEwen et al., 2015). Chronic cortisol elevations impair neurogenesis and synaptic plasticity in the hippocampus and PFC, which are critical regions for WM and memory consolidation. Cumulatively, the changes are manifested as persistent cognitive impairments, increased risk for depression and anxiety, and impaired adaptive functioning.
3.3 Developmental and Individual Differences in Stress-Working Memory Dynamics
The relationship between stress and WM is strongly guided by developmental stage, individual stress reactivity profiles, and early life experience. Tsai et al. (2021) demonstrated that older adolescents (13–15 years) derive greater WM benefits from acute coordinated stress responses than younger children (8–10 years), as this reflects increases in the maturational pattern of HPA-SNS regulatory circuits and PFC functioning. Children’s dysregulated or inconsistent neuroendocrine reactions at younger ages were associated with lower WM performance under stress, indicative of greater vulnerability at earlier developmental stages.
Besides, individuals who have been subjected to early adversity or maltreatment tend to exhibit dysregulated cortisol and catecholamine profiles, which make them prone to chronic stress states and consequent WM impairments (McEwen et al., 2015). Epigenetic mechanisms are likely responsible for these processes whereby early-life stress may affect the expression of genes regulating dopamine and norepinephrine receptors in the PFC and thus stress sensitivity and executive function outcomes (Chaby et al., 2015). Pilot data further suggest gender differences in stress reactivity and WM influence, but more research must be done to establish hormonal and genetic influences.
4. Discussion
The collective evidence characterizes the dualistic impact of stress on adolescent working memory that is sensitively mediated by neuroendocrine dynamics and developmental maturation. Acute stress, when defined by coordinated activation of the HPA and SNS, transiently improves WM by optimizing PFC neurotransmission through balanced catecholamine and glucocorticoid signaling. The cognitive facilitation is time-limited and dose-dependent, with variability thresholds for individuals and developmental stages. Conversely, chronic or dysregulated stress results in neuroplastic damage—dendritic retraction, synaptic loss, volumetric reductions—that impair PFC and hippocampal integrity and result in enduring WM impairments exacerbated by repeated stress exposure. Adolescent development is both a window of opportunity and vulnerability: system maturation enhances adaptive response capacity but also elevates risk for long-term cognitive sequelae if stress is prolonged or intense.
4.1 Education and Mental Health Intervention Implications
The marked WM impairments seen with chronic stress have significant implications for the quality of adolescent academic functioning and emotional health, highlighting the importance of early identification and intervention. Biomarkers such as salivary cortisol and alpha-amylase offer the noninvasive capacity to detect maladaptive patterns of stress response and enable timely psychosocial intervention. Mindfulness-based stress reduction, cognitive-behavioral therapy, and regular aerobic
exercise have supporting evidence for normalizing neuroendocrine function and facilitating WM. Moreover, pharmacological modulation of catecholamine and glucocorticoid receptors is a new field for the optimization of executive function in disorders of cognitive stress. Stage- and individual-specific interventions may optimally maximize cognitive resilience and long-term well-being.
5. Conclusion
Adolescence is a high neurodevelopmental stage where stress response and working memory dynamically feed back into one another, with long-term effects on emotional and cognitive processing. Acute stress, when characterized by balanced activation of the hypothalamic-pituitary-adrenal (HPA) axis and sympathetic nervous system (SNS), can temporarily enhance working memory via peak neurotransmitter signaling in the prefrontal cortex (Tsai et al., 2021; Arnsten, 2009). This adaptive response enables adolescents to develop executive control and cognitive flexibility under tolerable stress. However, this facilitation only depends on the precise timing and intensity of stress; disruptions in this exact balance are able to instantly switch acute stress from beneficial to detrimental (Shields et al., 2013).
In contrast, dysregulated or chronic stress during adolescence creates structural and functional alterations in brain regions involved in working memory, namely dendritic retraction, and hippocampal and prefrontal cortical volume loss (Lupien et al., 2009; McEwen et al., 2015). These neuroanatomical alterations equate to chronic deficits in working memory functioning and heightened risk for subsequent cognitive and emotional dysfunction. Importantly, developmental stage and stress reactivity interindividual differences moderate these effects, so that adolescents who are younger and who have experienced early-life adversity are at increased risk (Tsai et al., 2021; Chaby et al., 2015). Recognition of these subtle effects highlights the importance of early identification and specially targeted interventions—such as mindfulness, cognitive-behavioral therapies, and physical activity—that have the potential to rebalance stress systems and enhance cognitive resilience during this period of sensitivity.
Recognition of the dualistic impact of stress on the working memory of adolescents underscores the vulnerability and malleability of the developing brain. Additional integrative research combining neuroendocrine assessment, longitudinal neuroimaging, and behavioral testing is essential to clarify mechanisms and optimize interventions (McEwen et al., 2015; Tsai et al., 2021). These methods will play a central role in promoting cognitive well-being and emotional health in youth confronting the complex demands of adolescence and into young adulthood.
Acknowledgments
The writer is also keen on expressing their sincere appreciation to researchers whose early work provided an explanation of the intricate connection between stress and working memory in adolescents. Acknowledgement is also due to colleagues and mentors for useful comments and criticisms that shaped this review.
Works Cited
Arnsten, A. F. T. (2009). Stress signalling pathways that impair prefrontal cortex structure and function. Nature Reviews Neuroscience, 10(6), 410–422.
https://doi.org/10.1038/nrn2648
Chaby, L. E., Sheriff, M. J., Hirrlinger, A. M., & Braithwaite, V. A. (2015). Chronic unpredictable stress during adolescence impairs and improves learning and memory in adulthood. Behavioural Brain Research, 293, 168-175.
https://doi.org/10.1016/j.bbr.2015.07.014
Lupien, S. J., McEwen, B. S., Gunnar, M. R., & Heim, C. (2009). Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nature Reviews Neuroscience, 10(6), 434–445.
https://doi.org/10.1038/nrn2639
McEwen, B. S., Nasca, C., & Gray, J. D. (2015). Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology, 41(1), 3–23. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4675857/
Shields, G. S., Sazma, M. A., & Yonelinas, A. P. (2013). The effects of acute stress on episodic memory: A meta-analysis and integrative review. Psychological Bulletin, 139(6), 909–932.
https://doi.org/10.1037/a0035375
Tsai, J. L., Mukhopadhyay, S., & Quas, J. A. (2021). Acute stress-induced working memory changes in children: The role of sympathetic and HPA activation.
Psychoneuroendocrinology, 134, 105427.

Leave a comment